")
 Collared Flycatcher (Ficedula albicollis) is type of bird with limited area expansion. Their nesting enviroment is mostly older deciduous forests with enough sufficient of cavities in place 1200 – 1300 m above sea level.Article presents monitoring of nesting population from seven parts of Western Slovakia where in 2001 – 2011 we registered 329 nesting of this species bird in cabs. The moste filled cabs were in beech stand (max. 60%), at least orchard (10%). Average was 6.09 eggs after laying eggs on a whole found laying. Secondary production was set to 4.25 nestling and tertiary production in 3.94 nestling per nesting. The most common cause of nest loosing were Edible Dormouse (Glis glis) - 27% and Hazel Dormouse (Muscardinius avellanarius) - 20%. During the reporting period was marked 1399 individuals (including 1,286 nestling) and 27 controls reported captures.
Collared Flycatcher (Ficedula albicollis) is type of bird with limited area expansion. Their nesting enviroment is mostly older deciduous forests with enough sufficient of cavities in place 1200 – 1300 m above sea level.Article presents monitoring of nesting population from seven parts of Western Slovakia where in 2001 – 2011 we registered 329 nesting of this species bird in cabs. The moste filled cabs were in beech stand (max. 60%), at least orchard (10%). Average was 6.09 eggs after laying eggs on a whole found laying. Secondary production was set to 4.25 nestling and tertiary production in 3.94 nestling per nesting. The most common cause of nest loosing were Edible Dormouse (Glis glis) - 27% and Hazel Dormouse (Muscardinius avellanarius) - 20%. During the reporting period was marked 1399 individuals (including 1,286 nestling) and 27 controls reported captures.
1 Úvod
Muchárik bielokrký (Ficedula albicollis) má pomerne obmedzený typ rozšírenia. Areál druhu je súvislý od východného Francúzska, resp. južného Nemecka na východ až po Ukrajinu a juhozápadné Rusko. Izolované hniezdne populácie sa vyskytujú aj na juhu Apeninského polostrova ako aj na Baltských ostrovoch (Gotland, Őland - Švédsko) (Beklová & Formánek in Hudec, 1983). V 20-tom storočí sa druh rozšíril smerom na západ, avšak v súčasnosti sú západoeurópske populácie na ústupe. Kľúčové hniezdne populácie v oblasti rozšírenia (Rumunsko, Rusko, Ukrajina) sú hodnotené ako stabilné, alebo s rastúcim populačným trendom (BirdLife International, 2004). Slovensko je podľa EBCC atlasu domovom druhej najpočetnejšej populácie v Európe (podobne ako pri druhu Ficedula parva). Hniezdi výlučne v dutinách, či už prírodných, alebo umelých vtáčích búdkach. Sťahovavý druh, stredoeurópske populácie migrujú južnou alebo juhovýchodnou migračnou cestou do zimovísk vo východnej alebo južnej Afrike (Konžská Demokratická Republika, Uganda, Zambia) (Cepák et al., 2008). Na zimoviskách je typických habitatov riedky savanový les tzv. „miombo“. Migrácia má veľmi skoré načasovanie (už v priebehu júla), prílet na hniezdisko prebieha v apríli a máji (Snow & Perrins, 1998). Druh sa na hniezdisku zdržuje vôbec najkratšiu dobu zo všetkých našich nidifikantov (Cepák et al., 2008). V styčných územiach s muchárikom čiernohlavým (Ficedula hypoleuca) sa vyskytujú pravidelne zmiešané páry s plodným potomstvom (Beklová & Formánek in Hudec, 1983).

2 Metodika
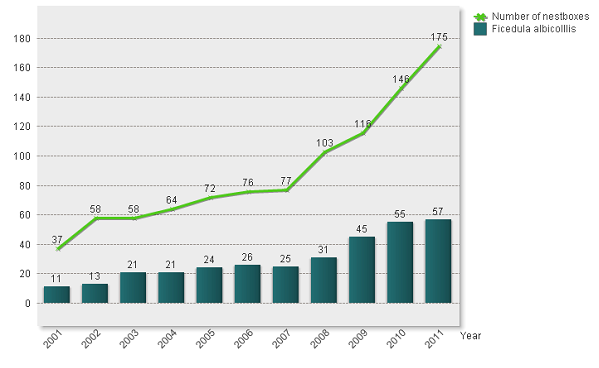
V rokoch 2001 – 2011 bol monitoring záujmového druhu založený na kontrolách vtáčích búdok, ktoré boli inštalované na siedmych plochách na šiestich lokalitách na jednej lokalite boli búdky situované do dvoch rozdielnych typoch porastu) v troch orografických celkoch západného Slovenska (okresy Prievidza, Bánovce nad Bebravou). Búdky boli inštalované s cieľom zistiť početnosť druhu v rozdielnych typov lesných porastov (viď. kap. 3). Búdky boli vyrobené podľa Zasadila (2001), pričom v zmysle tejto práce predstavovali tri základné typy – malý sýkorník, sýkorník a škorečník. Základný rozdiel v jednotlivých typoch je okrem vonkajších rozmerov vletový otvor búdky (2,8 cm, 3,2 cm, 5,0 cm). Počet búdok v jednotlivých rokoch bol rôzny, pričom mal stúpajúcu tendenciu (Graf 1).
Monitoring priebehu hniezdenia druhu prebiehal od poslednej pentády apríla až do ukončenia hniezdneho procesu posledného hniezdiaceho páru v každom roku, pričom búdky boli kontrolované pravidelne každú pentádu. V rámci hniezdneho procesu boli sledované základné charakteristiky v zmysle práce Országhová & Blazníková (1998): primárna produkcia (počet vajec v kompletnej znáške), sekundárna produkcia (počet vyliahnutých mláďat), terciálna produkcia (počet vyletených mláďat). Zisťovali sme aj fidelitu a filopatriu a to pomocou odchytu a značenia inkubujúcich samíc sa hniezde a krúžkovania mláďat. Mláďatá boli najčastejšie krúžkované medzi 5-tym až 10-tym dňom od narodenia, aby sa zamedzilo predčasnému opusteniu hniezda. V jednom prípade pre vysoký vek mláďat bolo upustené od ich značenia (rok 2009).
3 Charakteristika štúdijných plôch
Hniezdenie muchárika bielokrkého bolo sledované na siedmych záujmových lokalitách, ktoré predstavovali rôzne lesné typy porastov z hľadiska veku ako aj štruktúry (podrobne uvedené pri opise lokalít) a jeden nelesný typ – ovocný sad. Monitoring prebiehal na území dvoch okresov – Prievidza a Bánovce nad Bebravou, v troch orografických celkoch: v pohoriach Strážovské vrchy (kód orografického celku 120): lokalita č.1, 2, 6 a 7), Vtáčnik (280): lokalita č.4 a 5, a v Hornonitrianskej kotline(230): lokalita č.3).
Lokalita č.1: k.ú Chvojnica, okres Prievidza - 60 ročný bukový porast v úvode Hluchej doliny. Prímes tvoria čerešňa vtáčia (Cereasium avium), borovica lesná (Pinus sylvestris). Počet búdok v roku 2001 - 9, v roku 2011 - 4. Súradnice, stred lokality: 48° 52´ N, 18° 33´ E.
Lokalita č.2: k.ú Nitrianske Pravno, miestna časť Bôrik, okres Prievidza - 40 ročný extenzívne obhospodarovaný jabloňový sad (Malus domestica). Počet búdok v roku 2011 -10. Súradnice, stred lokality: 48° 52´ N, 18° 37´ E
Lokalita č.3: k.ú Prievidza, okres Prievidza - 50 ročný sekundárne zmiešaný lesný porast v blízkosti sedimentačnej jamy Bane Cígeľ s dominantným zastúpením borovice lesnej (Pinus sylvestris), dubu zimného (Quercus petraea) a smreka obyčajného (Picea abies). Počet búdok v roku 2005 – 9, v roku 2011 – 14. Súradnice, stred lokality: 48° 45´ N, 18° 38´ E
Lokalita č.4: k.ú Kamenec p. Vtáčnikom, okres Prievidza - okraj 50 ročného dubovo - hrabového porastu Bystričianskej doliny a úvode Gepnárovej doliny na pravom brehu potoka. Prímes tvorí čerešňa vtáčia (Cereasium avium) a buk lesný (Fagus sylvatica). Podrast je zastúpený miestami najmä lieskou obyčajnou (Corylus avellana). Počet búdok v roku 2001 - 12, v roku 2011 - 62. Súradnice, stred lokality: 48° 39´ N, 18° 34´ E
Lokalita č.5: k.ú Kamenec p. Vtáčnikom, okres Prievidza - 50 ročný bukový porast v Gepnárovej doline na ľavom brehu potoka. Prímes tvorí smrek obyčajný (Picea abies) a hrab obyčajný (Carpinus betulus).Počet búdok v roku 2001 - 16, v roku 2011 - 45. Súradnice, stred lokality: 48° 39´ N, 18° 35´ E
Lokalita č. 6: k.ú Látkovce, okres Bánovce nad Bebravou - okraj 40 ročného monotypického dubového porastu na pravo od štátnej cesty Dolné Vestenice - Jankov Vŕšok. Počet búdok konštantný od roku 2008: 20 Súradnice, stred lokality: 48° 43´ N, 18° 21´ E
Lokalita č. 7: k.ú Uhrovské Podhradie, okres Bánovce nad Bebravou - 30 ročný monotypický bukový porast v doline západne od vrcholu Rokoša.. Počet búdok konštantný od roku 2010: 20. Súradnice, stred lokality: 48° 46´ N, 18° 25´ E
)")
4 Výsledky
Počas rokov 2001-2011 bolo registrovaných 329 hniezdení muchárika bielokrkého v sledovaných lokalitách. Najmenej – 11 hniezdení bolo v roku 2001, najviac 57 v roku 2011, čo je výsledkom vyššieho počtu inštalovaných búdok (Graf 1). Z hľadiska typu búdky boli najčastejšie využívané pre hniezdenie sýkorníky s vletovým otvorom 3,2 cm – 256 prípadov (78%). 64 krát (19%) využili pre hniezdenie mucháriky búdky typu škorečník a len v 9 tich prípadoch malý sýkorník (3%).
Na jednotlivých sledovaných lokalitách bolo zastúpenie záujmového druhu rôzne. Početne najvýznamnejšie hodnotíme zastúpenie druhu v búdkach v bukových porastoch (lokality č.1, 5 a 7). Obsadenie v búdkach na lokalite č.1 oscilovalo medzi 0 – 5 pármi/rok, čo predstavovalo maximálnu obsadenosť 55,56% (rok 2004). Vysokú amplitúdu možno pripisovať pomerne nízkemu počtu búdok (4 - 9 kusov), čím je obmedzená reprezentatívnosť získaného vzorku. Na lokalite č. 5 sa pohybovala obsadenosť búdok v rozmedzí od 25 (rok 2001) do 60% (roky 2006 a 2009), pričom po roku 2005 neklesla pod hranicu 40%. Tretia lokalita predstavujúca bukový porast je charakteristická svojou konštantnosťou po oba sledované roky, kedy obsadenosť predstavovala – 40%. Dubo-hrabové porasty reprezentujúce lokalitu č. 4 sú druhom pravidelne, aj keď menej početne obsadzované. Obsadenosť búdok v takomto type porastu predstavovalo rozmedzie od 18,18 do 36,36%. Maximálny počet párov sme registrovali v roku 2011 – 21, čo je dôsledkom vyššieho počtu búdok ako v predchádzajúcich sezónach – 62. Lokalitu č. 6 reprezentujúcu dubový porast charakterizuje 5 – 9 hniezdení v rámci jednej hniezdnej sezóny, s minimom v roku 2008 a maximom v roku 2009. Obsadenosť v rámci jednej hniezdnej sezóny činí od 25 do 40%. Pomerne nejasná situácia je v zmiešanom type porastu (lokalita č. 3), kde sme v rokoch 2006-2009 nezaznamenali ani jedno hniezdenie, ale v roku v roku 2009 až 3 hniezdenia. Obsadenosť búdok v rámci jednej hniezdnej sezóny preto osciluje medzi 0 až 21,43%. Reálnejší obraz by poskytol vyšší počet búdok, keďže na lokalite bolo v sledovanom období monitorovaných 9 – 14 búdok. Najnižšie hodnoty boli dosiahnuté v záhradách, kedy na lokalite č. 2 reprezentuje bola obsadenosť 10%, čo však vzhľadom na počet búdok reprezentuje len jedno hniezdenie druhu.
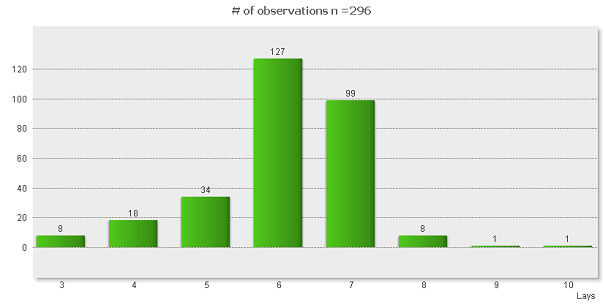
V 296-tich prípadoch došlo k zneseniu úplnej znášky (1804 vajec), pričom 33-krát došlo k predácii hniezda ešte počas znášania vajec (10,03%). Priemerne bolo znesených 6,08 vajec na jednu kompletnú znášku. Najvyššia priemerná bola zaznamenaná v roku 2008 – 6,79 vajec, najnižšia v roku 2004 – 5,58 vajec. Najpočetnejšia bola znáška o počte o šiestich vajec – 127 prípadov (42,91%). Najmenej zniesli mucháriky 3 vajcia – 8 prípadov (2,70%). Najväčšia znáška bola zistená o desiatich vajec – jeden prípad (lokalita č.4, 2008) a jedenkrát bola zistená aj deväťpočetná násada (lokalita č.5, 2008, Graf 2).
Terciálnu produkciu tvorí 1297 mláďat, ktoré vyleteli z búdok. Aspoň jedno mláďa vyletelo v 240-tich prípadoch (72,95%). Priemerne vyletelo 3,94 mláďaťa/1 hniezdo (n = 329). Najvyššia bola v rokoch 2007 - 4,8 a v 2011 - 4,75, najnižšia v 2010 – 2,76 a 2004 – 2,86 mláďať/1 hniezdo. Najviac mláďat z jedného hniezda - 8 vyletelo v dvoch prípadoch. Najmenej – 1 v piatich prípadoch (Graf 3).


Dostatočný dátový súbor z lokalít č.4 a 5 umožňuje vzájomnú komparáciu dosiahnutých výsledkov. Primárna produkcia dosahuje priemerné vyššie hodnoty v bukovom poraste v 9tich z monitorovaných 11tich sezón (mimo rokov 2002 a 2006).V prípade sekundárnej sukcesie sú hodnoty z oboch typov porastov vyrovnanejšie, nakoľko vyššia sekundárna produkcia bola v búdkach bučín zaznamenaná počas siedmych hniezdnych sezón (2003, 2004, 2006, 2007, 2009, 2010 a 2011). Naviac situáciu v rokoch 2001 a 2002 môže skreslovať nižší počet inštalovaných búdok a z toho vyplývajúca nižšia interpretovateľná hodnota dosiahnutých výsledkov. Z hľadiska dosiahnutých hodnôt terciálnej produkcie je situácia obdobná u sekundárnej.
Predácia bola zaznamenaná v 89tich prípadoch hniezdenia (27,05%). Z tohto počtu tvorí 27% predácia plchom veľkým (Glis glis). Pätinu zo všetkých predovaných hniezd registrujeme u pĺšika lieskového (Muscardinius avellanarius). Tretí najčastejší prípad neúspešného hniezdenia je ulovenie samice, čoho prejavom bolo nájdenie uhynutých mláďat, alebo opustenie oplodnenej znášky v čase inkubácie. U krutihlava hnedého (Jynx torquilla) evidujeme 4 prípady likvidácie hniezdiska muchárika. V týchto prípadoch nešlo o predáciu hniezda z trofických dôvodov, ale z dôvodu záujmu o dutinu – búdku z dôvodu výberu daného stanovišťa za svoje vlastné hniezdisko (Graf 5).


5 Diskusia
V priebehu jedenástich hniezdnych sezón bol získaný dátový súbor z 329 hniezdení muchárika bielokrkého. Z hypsometrického pohľadu bola populácia monitorovaná v lokalitách, kde dochádza k pomerne vysokej koncentrácii druhu v rámci Slovenska, nakoľko Kropil et al. (1995) udáva priamu závislosť druhu na listnatých porastoch. Krištín (1991) konštatuje z územia Poľany najvyššie populácie v dubovo-hrabových porastoch (3,6 páru/10 ha), v bukovo-jedľových porastoch už len 2 páry/10 ha, pričom v horských smrečinách druh úplne absentuje. Z Vtáčnika Krištín & Harvančik (1992) udávajú druh ako hniezdiča až vo výškach v 1200 – 1300 m n. m. V podmienkach lužného lesa je druh ešte bežnejší. Bohuš et al. (1999) zistil vo vŕbovo-topoľovom lužnom lese v NPR Ostrov orliaka morského 4-13 párov/10ha. Korňan (1996) udáva z jelšového lužného lesa 9,4-11,3 páru/10 ha a z topoľového lužného lesa 6,4-6,7 páru/10 ha. Vysoké hodnoty konštatuje Kropil (1993) z dubín so slabo vyvinutým podrastom z územia Tríbeča: 8,9-12,8 páru/10ha, resp. 14,4-21,3páru/10 ha. Ten istý autor konštatuje pokles denzity druhu v hospodárskych porastoch, čo je logické vzhľadom na absenciu dutín, ktorých výskyt je nevyhnutný predpoklad pre zahniezdenie druhu. Z mestského prostredia mestských parkov a záhrad zo Zvolena ho udáva Krištín & Zach (1993), resp. Műllerová (1996). Tu hustota dosahuje 3,3-3,9 páru/10 ha. Krištín & Kropil in Danko et al. (2002) upozorňujú na význam búdok z hľadiska praktickej ochrany druhu a udávajú v takýchto podmienkach hustotu nad 20párov/10 ha.
 |
|
 |
Podobne v Českej republike hniezdi najmä v nižšie položených lesoch. Na území dochádza naviac k poklesu počtu druhu smerom z juhovýchodu na severozápad. Hypsometricky dosahuje najvyššie položené hniezdisko v oblasti Velké Javoriny v Bílych Karpatech (Šťastný et al., 2006), v Moravsko-Slezských Beskydech je to potom do 900 m n.m (Čapek, 1994). Efekt vyvesovania vtáčích búdok na zvýšenie populácie druhu sledoval Král (1991a) na lokalite Sovinec v Jeseníku, ktorý uvádza 1,11-4,67 samice/1 ha. Autor upozorňuje na opatrnosť interpretácie počtu párov na plošnú jednotku, nakoľko samce bývajú u tohto druhu často polygamné (Lőhrl, 1949; Král, 1991 b).
Z hľadiska počtu znášky je celkový priemer na nami získaných lokalitách v rámci sledovaného obdobia 6,09 vajca/1 hniezdenie. Najvyššiu sezónnu/ročnú hodnotu sledujeme v roku 2008 - 6,79, pričom najvyššia hodnota na lokálnej úrovni pripadá na rok 2008 na lokalitu č.5 – 6,85. Tieto hodnoty sú porovnateľné s tými, ktoré udávajú Král & Krause (2010) z oblasti lužného lesa v blízkosti Břeclavi. Autori konštatujú najvyššiu sezónnu primárnu produkciu 6,87 vajca s dlhodobým priemerom (1988-2009) 6,57 vajca/1 hniezdenie. Ďalej v oblasti Nízkeho Jeseníku bola najvyššia priemerná sezónna primárna produkcia 6,69 vajca s dlhodobým priemerom 6,30 vajca/1 hniezdenie. Stříteský (2010) udáva z dubového porastu (nadmorská výška 420-430 m n. m) pre roky 2005-2010 priemerne 6,12 vajca/1 hniezdenie s maximom v roku 2008-6,33 a minimom v roku 2010-5,62 vajca/1 hniezdenie. Hudec (1983) uvádza v rámci celej Českej republiky priemerne 5,63 vajca/1 hniezdenie, pričom v práci bola maximálne zistená znáška o počte 8mich vajec, pričom naše zistenia predkladajú jednu 9 početnú znášku a jednu o počte 10 vajec. Géroudet (1958) udáva sedenie samice od predposledného vajca, pričom Balát (1976) konštatuje súčasné liahnutie mláďat, pričom naše zistenia tomuto údaju v mnohých prípadoch odporujú, kedy na mláďatách v mnohých prípadoch bol viditeľný rozdiel vo veku aj 10 dní po vyliahnutí najstaršieho jedinca, čoho dôkazom je aj váha mláďat, ktoré boli vážené na lokalitách číslo 3 a 6. Tu bol rozdiel medzi najmladším a najstarším jedincom 1,5g (lokalita č.3), resp. 4g (lokalita č.6)!

Extrémne fenologické údaje charakterizujú dva prípady hniezdenia v roku 2007, kedy boli 14.5.2007 v dvoch búdkach krúžkované mláďatá vo veku 7, resp. 9 dní (lokality č. 4 a 5). Pri započítaní doby inkubácie 13 - 15 dní, doby znášania vajec (každý deň znesie samica jedno vajce) a stavby hniezda 5 dní dochádzame k záveru, že s hniezdením začali oba páry medzi 13.4. - 15.4.2007 a znesenie prvého vajca medzi 18.4. - 20.4.2007. Táto hodnota koreluje s výsledkom z Nízkeho Jeseníku (lokalita Dolná Součka - Sovinec), kedy bol zaznamenaný prílet samca na lokalitu už 9.4.2009 a dátum znesenia prvého vajca bol 20.4.2009 (Král & Krause, 2010). Z lužných lesov lokality Břeclav – Pohansko dokonca autori udávajú dátum znesenia prvého vajca už z 13.4.2009. Tieto extrémne hodnoty dávajú autori do súvisu s teplým počasím, ktoré bolo sprievodným javom v priebehu marca a apríla v oblasti juhovýchodnej migračnej cesty, ktorú druh využíva.


Straty na hniezdach košntatuje Hudec (1983) na 28,8%., čo je veľmi podobná hodnota dosiahnutá na našich lokalitách – 27,05%. Autor spomína ako hlavný druh predátora kunu lesnú (Martes martes) a ďatľa veľkého (Dendrocopos major). Henze (1943) spomína ako možného predátora druhu aj ryšavky (Apodemus sp.), Glowaciński (1973) lasicu, pričom ani jeden zo spomínaných druhov sme my ako predátora hniezda nezistili. Stříteský (2010) pripisuje straty na hniezdach plchovi veľkému (Glis glis) a nepriaznivým poveternostným podmienkam. Hniezdna úspešnosť v rokoch 2005-2010 oscilovala medzi 32-74%, s minimom v roku 2005 a maximom v roku 2007. Adamík & Král (2008) porovnávajú z Moravských hniezdísk predátorov dutinových hniezdičov tri druhy plchov – Glis glis, Dryomis nitedula a Muscardinius avellanarius. Najčastejšie sa v sledovanom období (1975 – 2005) na predácii podieľal Glis glis– 331 predovaných hniezd, ďalej Muscardinius avellanarius – 59 prípadov a Dryomis nitedula – 57 prípadov, pričom najčastejšie predovaným druhom bol práve Ficedula albicollis. Celkové straty boli stanovené na 18,44%. V Nemecku Koppmann-Rumpf et al. (2003) bola vyčíslená predácia u príbuzného Ficedula hypoleuca (Ficedula albicollis sa v Nemecku vyskytuje ojedinele) na 29,36%, v Litve potom Juškaitis (2006) na 17,4%. Závislosť medzi zvýšenou predáciou cicavcami a oteplovaním klímy konsťatujú Adamík & Král (2007).

Celkovo bolo v rokoch 2001 - 2009 na Slovensku okrúžkovaných 1749 jedincov, z toho 1371 mláďat (Slobodník, 2010). Na nami monitorovaných lokalitách to bolo do roku 2009 928 jedincov, z toho 867 mláďat na hniezdach. Spomínaná hodnota tak predstavuje 53,06% podiel z celoslovenského počtu okrúžkovaných jedincov, čím je významná nie len z hľadiska lokálneho študia populácie, resp. nidobiológie druhu, ale má aj celonárodný význam z hľadiska monitoringu špecializovanej krúžkovateľskej činnosti. Valeš (2009) spomína zahájenie monitoringu populácie muchárika bielokrkého v Českej republike pomocou krúžkovania metodikou RAS (Retraping Adult for Survival), ktorý kontinuálne nasledoval aj v roku 2010 (Valeš, 2010). Základom výskumu bol monitoring 105tich búdok v dubových porastoch Vojenského újezdu Březina, kde bolo sledovaných na ploche 14 ha po obe sezóny 34 hniezdení druhu. Z hľadiska dlhovekosti patrí nami získaný údaj medzi najstaršie jedince v rámci Slovenska. Jedinec bol v čase kontroly v piatom roku života. Staav & Fransson (2006) udávajú vo Švédsku kontrolný odchyt jedinca, ktorý sa dožil veku 7 rokov a 11 mesiacov. Adamík & Král (2007) kontrolovali v roku 2005 jedinca, ktorý bol označený 25.5.1998 ako samec minimálne v treťom roku života, čo znamená, že v čase kontroly mal deväť rokov! Neoficiálne je však najstarším jedincom samica krúžkovaná súkromným krúžkom na ostrove Gotland vo Švédsku (Staav in verb).
5 Záver
V priebehu sledovaného obdobia (2001 - 2011) sme monitorovali 329 prípadov hniezdenia muchárika bielokrkého (Ficedula albicollis) na siedmych lokalitách, v rozličných typoch lesného porastu a jedného nelesného. Najvyššie hodnoty obsadenosti búdok sme zaznamenali v bukových porastoch, stredné v dubovo-hrabových a dubových porastoch. Najmenej osídlené boli búdky v zmiešaných porastoch a ovocnom sade, kde došlo k zahniezdeniu iba jedného páru. Priemerná primárna produkcia bola na hniezdach s úplnou násadou 6,09 vajec/1 hniezdenie (n = 296). Sekundárna 4,25 vyliahnutých mláďat/1 hniezdo a terciálna 3,94 vyletených mláďat/1 hniezdo (n = 329). Na hniezdnych stratách sa zo známych pričín podieľali najviac plch veľký (Glis glis) a pĺšik lieskový (Muscardinius avellanarius). Spolu bolo okrúžkovaných 1399 jedincov (z toho 1296 mláďat), pričom registrujeme 27 kontrolných odchytov potvrdzujúcich vysoku fidelitu a filopatriu druhu. Komunikáciu medzi vzdialenejšími lokalitmi (presun na väčšiu vzdialenosť) sme nezaznamenali.
6 References
Adamík, P. & Král, M. 2007. Záznam neobvykle starého kroužkovance lejska bělokrkého (Ficedula albicollis) na Moravě. Kroužkovatel 3: 20.
Adamík, P. & Král, M., 2008: Nest losses of cavity nesting birds caused by dormice (Gliridae, Rodentia). Acta Teriologica 53 (2): 185-192.
Adamík, P. & Král, M., 2007: Climate- and resource-driven long-term changes in dormice populations negatively affect hole-nesting songbirds. Journal of Zoology 275: 209–21
Balát, F., 1976: Fortpflanzungsökologie der höhlenbrűtenden Vögel in sűdmährischen Kieferwald. Acta Sc. Nat. Brno 10: 1-44
BirdLife International, 2004: Birds in Europe: Population estimates, trends and conservations status. Cambridge, UK, Birdlife Consevations Series No. 12
Bohuš, M., Baloghová, A., Illavský, J. & Kalúsová, E., 1999: Príspevok k poznaniu hniezdnych ornitocenóz vybraných lesných porastov inundačného územia Dunaja. Tichodroma 12: 61-91.
Čapek, M., 1994: Birds in mountain ecosystems under pressure of air pollution. Acta Sc. Nat. Brno 28: 1-46
Danko, Š., Darolová, A., & Krištín., A., 2002: Rozšírenie vtákov na Slovensku. Veda, Bratislava, 686 s.
Géroudet, J., 1961: Les passereaux. I (2. éd.). Paris
Glowacinski, Z., 1973: Phenology and breeding success in a population of Collared Flycatcher, Ficedula albicollis (Temm.), in the Niepołomice Forest (Southern Poland).
Henze, O., 1943: Vogelschutz gegen Insektenschaden in der Fortwirtschaft. Műnchen.
Hudec, K. et al., 1983: Fauna ČSSR. Ptáci 3/I,II. Academia, Praha, 1236 s.
Juškaitis R. 2006: Interactions between dormice (Gliridae) and hole-nesting birds in nestboxes. Folia Zoologica 55: 225–236.
Koppmann-Rumpf B., Heberer C. and Schmidt K. H., 2003: Long-term study of the reaction of the edible dormouse Glis glis to climatic changes and its interactions with hole-breeding passerines. Acta Zoologica Academiae Scientiarum Hungaricae 49 (Suppl. 1): 69–76.
Korňan, M., 1996: Analýza štruktúry ornitocenózy nížinných lesov na Slovensku a vplyv migračných gíld na formovanie lesných ornitocenóz v závislosti od výškového gradientu. Diplomová práca, Prírodovedecká fakulta UK, Bratislava, 114 s.
Král M., 1991: Polyteritoriální chovaní a sukcesivní polygamie samců lejska bělokrkého (Ficedula albicollis Temm.) v Nízkem Jeseníku. Panurus 3: 159-168.
Král M., & Krause F., 1991: Charakteristika lužní a submontánní populace lejska bělokrkého (Ficedula albicollis Temm.) na Moravě. Zprávy MOS 49: 37–44.
Král M., & Krause F.,2010: Extrémní fenologická data lejska bělokrkého (Ficedula albicollis) ve dvou biotopech na Moravě. Sylvia 46: 63-69.
Krištín, A. & Harvančík, S., 1992: K štruktúre a ekologiíí vtáctva na Vtáčniku. - Rosalia 9: 223–232.
Krištín, A. & Zach, P., 1993:Ornitocenózy južného Podpoľania navrhovaného významného vtáčieho územia Európy: 106-116. In: Urban, P., (ed.), Fauna Poľany, MaB, Zvolen.
Kropil, R., 1993: Štruktúra a produkcia ornitocenóz vybraných prírodných lesov Slovenska. Kandidátska dizertačná práca, Lesnícka fakulta TU, Zvolen, 129 s.
Kropil, R., Dudich, A. & Finďo, S., 1995: Natural forests of the western Carpathians – important areas of animal diversity: 55-61. In: Paulenka, J & Paule, L.: Conservations of forests in Central Europe, Arbosa Publishers, Zvolen.
Műllerová, M., 1996: Štruktúra a dynamika hniezdnej ornitocenózy parku v Rusovciach. Tichodroma, Bratislava, 9, str. 73-79
Löhrl, H., 1949: Polygamy among Collared Flycather (Ficedula albicollis). Vogelwarte: 94 Országhová, Z., Blázniková, V., 1998: Hniezdenie sýkoriek (Parus major, Parus caeruleus) v búdkach v Botanickej záhrade Univerzity Komenského v Bratislave. Tichodroma, 11:118-132.
Snow, D.W. & Perrins, C.M., 1998: The Birds of Western Paleartic. Oxford University Press, Oxford, 1653 s.
Slobodník, R., 2011: Zhodnotenie migračných trendov vtákov pomocou krúžkovateľských výsledkov na Slovensku. Rigorózna práca, Prírodovedecká fakulta UKF, Nitra.
Šťastný, K., Bejček, V. & Hudec, K., 2006: Atlas hnízdního rozšíření ptáků v České republice. Aventinum, Praha.463 s.
Staav R., & Fransson T., 2006: EURING list of longevity records for European birds. http://www.euring.org/data_and_codes/longevity.htm
Stříteský. J, 2010: Výzkumný projekt RAS – hnízdění lejska bělokrkého v umělých dutinách ve Vojenském újezdu Březina. Zprávy RAS 2: 5-8
Valeš. Z, 2010: Zprávy RAS Retraping Adults for Survival Opakovaný odchyt dospělců pro určení. Zprávy RAS 1.
Zasadil, P., 2001: Ptáčí budky a další zpúsoby zvyšovaní hnízdnych možností ptákú. Praha, 136